Part VI: The Biological Receiver
Chapter 14: The Synaptic Abyss and Macroscopic Coherence
14.1 The Humpty Dumpty Paradox (The Binding Problem)
In the late 19th and early 20th centuries, as neuroanatomists began to map the cerebral cortex, they discovered a strict division of labor within the human brain. The brain does not process reality in a centralized location. It compartmentalizes information.
Imagine you are standing in a forest, watching a bright red cardinal sing on a nearby branch. The optical data of the bird's shape and its red hue is routed from your retinas to the primary visual cortex (V1 and V4) at the back of your skull. The auditory data of the bird's song is routed from your cochlea to the temporal lobes on the sides of your head. The spatial awareness of where the bird is located relative to your own body is processed in the parietal lobe near the top of your brain. The emotional recognition of the moment is processed within the limbic system, while the semantic classification (identifying the bird as a "cardinal") is handled by the prefrontal cortex directly behind your forehead.
The physical data of that single moment is distributed across billions of neurons separated by macroscopic distances---inches of folded biological tissue.
And yet, you do not experience a fragmented reality.
You do not experience the color red in the back of your head, the sound of the song on the sides of your head, and the spatial location at the top of your head as disjointed, asynchronous data streams. You experience a single, unified moment of conscious awareness. The cardinal, its song, its location, and its color are bound together into one singular conscious object.
In classical neuroscience and the philosophy of mind, this paradox is known as the Binding Problem [1].
How does the brain put Humpty Dumpty back together again? How do 86 billion physically separated, chemically isolated neurons, firing at slightly different times and processing different modalities of sensory data, synchronize to create the illusion of a unified "Self"?
For decades, neuroscientists have approached this paradox using the tools of classical physics and chemistry. One prominent hypothesis is "gamma-band synchronization"---where distant groups of neurons fire their action potentials in rhythm (roughly 40 times a second), and this synchronized electrical activity weaves the disparate data together into a conscious whole.
But this classical explanation faces a physical constraint.
The transmission of information in the classical brain is relatively slow. When a neuron fires an action potential down its axon, the electrical spike travels at a maximum of roughly 100 meters per second. When it reaches the end of the axon, it must convert its electrical charge into a chemical payload (a neurotransmitter). Those chemicals must drift across the fluid-filled gap between the neurons (the synaptic cleft), bind to receptors on the other side, and trigger a new electrical spike. This entire process of synaptic transmission takes roughly 1 to 2 milliseconds per connection.
If the visual cortex at the back of the brain needs to synchronize with the prefrontal cortex at the front, the signal must cross millions of these chemical synapses. By the time the classical electrical signal travels across the brain, the external moment has passed. The processing speeds, vast distances, and variables of chemical diffusion struggle to account for the instantaneous unity of conscious experience.
Two separate metronomes ticking at the exact same speed on opposite sides of a room do not physically merge into a single, subjective entity. Synchronized syntax does not create unified semantics.
Classical physics, limited by the speed of chemical diffusion and the rigid boundaries of the cell membrane, struggles to fully resolve the Binding Problem.
To create a unified conscious experience across macroscopic distances, the framework proposes that the brain cannot rely solely on chemical messengers. It requires a physical mechanism that is non-local and mathematically capable of binding disparate physical locations into a single geometric state.
It requires Macroscopic Quantum Coherence.
But to achieve this, the quantum entanglement generated in Chapter 11 must break out of its cellular isolation. It must cross the abyss between the neurons.
14.2 The Synaptic Cleft: The Moat of the Mind
A defining architectural feature of the human brain is the empty space between its cells.
When Santiago Ramón y Cajal proved that the brain was not a continuous web of tissue (disproving the reticular theory), he revealed that every neuron in your brain is a physically isolated island. They reach out and touch each other with branching dendrites and axons, but they never fuse.
Between the axon terminal of one neuron (the sender) and the receiving dendrite of another (the receiver) lies a fluid-filled gap roughly 20 to 40 nanometers wide. This is the Synaptic Cleft.
For a classical electrical signal to continue its journey, it must stop at the edge of this gap, translate its electrical charge into a chemical payload, release those chemicals into the void, and wait for them to diffuse across to trigger the next neuron.
For the quantum entanglement locked inside our Posner molecules, the synaptic cleft is a biological gauntlet.
The intracellular fluid inside the neuron is carefully regulated. But the extracellular matrix filling the synaptic cleft is a chaotic environment. It is an ocean of roaming enzymes, voltage gradients, metabolic waste, and storms of calcium and sodium ions. If the quantum state of the phosphorus nuclei is to survive the journey between cells, it must remain armored, and its transit must be orchestrated.
If an entangled phosphorus atom attempts to cross this gap unprotected, the magnetic fields of the extracellular fluid will breach its electron cloud, disrupting the Singlet State. The entanglement will be lost to decoherence. The quantum bridge will collapse at the border of the cell.
To safely transport the quantum payload across the abyss, biology cannot simply release the atoms into the void. It must utilize a shielded transport mechanism. It must use a Trojan Horse.
14.3 The Trojan Horse Mechanism (Vesicular Exocytosis)
Let us zoom in on an axon terminal---the physical edge of a transmitting neuron bordering the synaptic cleft.
Inside this terminal, thousands of microscopic lipid bubbles are waiting. These are Synaptic Vesicles. In classical neuroscience, these vesicles act as transport containers, filled with signaling chemicals like glutamate, dopamine, or GABA.
In the framework of Dimensional Field Theory, drawing on the quantum biological models of Matthew Fisher, these vesicles carry a secondary payload [2].
As the Posner molecules () drift down the axon, carrying their entangled Phosphorus-31 nuclei, specialized transport proteins recognize their geometric shape. Because Posner molecules are small---roughly 0.87 nanometers in diameter---they are swept up and packaged inside these synaptic vesicles alongside the standard glutamate neurotransmitters.
The vesicle acts as a biological Trojan Horse. It is a sealed, lipid-bilayer transport. Inside, the classical neurotransmitters and the quantum-entangled Posner molecules float together, protected from the surrounding cytoplasm.
When the electrical action potential reaches the axon terminal, it triggers a mechanical cascade. Voltage-gated channels open, and an influx of calcium ions enters the cell. This calcium activates a winch system made of SNARE proteins. The SNARE proteins snare the lipid vesicles, draw them to the edge of the cell membrane, and zipper together.
In a process called Exocytosis, the lipid layer of the vesicle fuses with the lipid membrane of the neuron. The vesicle opens outward.
The payload is released into the synaptic cleft.
A flood of glutamate molecules crosses the 20-nanometer gap, binding to the NMDA and AMPA receptors on the post-synaptic neuron, triggering the classical electrical signal.
Swimming silently within that release of glutamate are the Posner molecules. They are expelled into the extracellular fluid. For a fraction of a millisecond, they are adrift in the moat.
14.4 Navigating the Void and Cellular Uptake
As the Posner molecules tumble through the synaptic cleft, they are subjected to the thermal and electrical noise of the extracellular environment. But the geometry of the Posner molecule holds firm.
The nine calcium ions forming the outer shell possess a nuclear spin of exactly zero. They are magnetically inert. As the molecule tumbles through the fluid, a quantum mechanical effect known as motional narrowing averages the external magnetic noise to zero. The Faraday cage holds. The Singlet State of the phosphorus nuclei inside remains isolated, maintaining quantum entanglement with its twin, which might currently reside millimeters away in the original brain region.
The journey takes merely a few microseconds.
On the other side of the gap lies the post-synaptic neuron. Its surface is studded with receptor proteins and ion channels reacting to the glutamate. But the receiving cell also possesses mechanisms to recover calcium and phosphate, essential building blocks for its own survival and energy metabolism.
Through transport channels, or through a bulk process called clathrin-mediated endocytosis (where the receiving cell membrane pinches inward to absorb extracellular fluid), the Posner molecules are drawn into the new neuron.
The crossing is complete. The quantum vaults have breached the neighboring cell.
Consider what has occurred. A Posner molecule generated in Neuron A is now located inside the cytoplasm of Neuron B. But this molecule still contains the entangled Phosphorus-31 nuclear spin whose twin remains inside Neuron A.
The two neurons, physically separated by the synaptic cleft and chemically isolated by their lipid membranes, are now quantum entangled. They share a single wave function that spans the void.
But for the brain to function as a unified entity, entangling two adjacent neurons is not enough. The network must scale. It must entangle billions of neurons across the volume of the cortex, connecting the visual cortex to the frontal lobe.
If every neuron had to physically swap a Posner molecule with every other neuron to achieve network entanglement, the process would take years. The logistics of moving molecules across inches of brain tissue are too slow to account for the instantaneous unity of consciousness.
To solve this logistical hurdle, the framework proposes that biology utilizes a specific protocol from quantum information theory.
14.5 The Quantum Viral Vector (Entanglement Swapping)
In 1993, physicists working in quantum teleportation and cryptography detailed a mechanism in the laws of entanglement. This protocol was demonstrated in a laboratory setting in 1998 by Anton Zeilinger (who later won the Nobel Prize for his work) [3].
They demonstrated a process that allows two quantum particles to become entangled even if they have never physically interacted.
This is Entanglement Swapping.
To understand this protocol, imagine four separate quantum particles: A, B, C, and D.
Suppose that Particle A and Particle B are generated together in Laboratory 1, and they are quantum entangled. They share a single wave function. In Laboratory 2, Particle C and Particle D are generated together, and they are also quantum entangled.
At this moment, A is entangled with B, and C is entangled with D. But A has no relationship with C or D. They are independent systems.
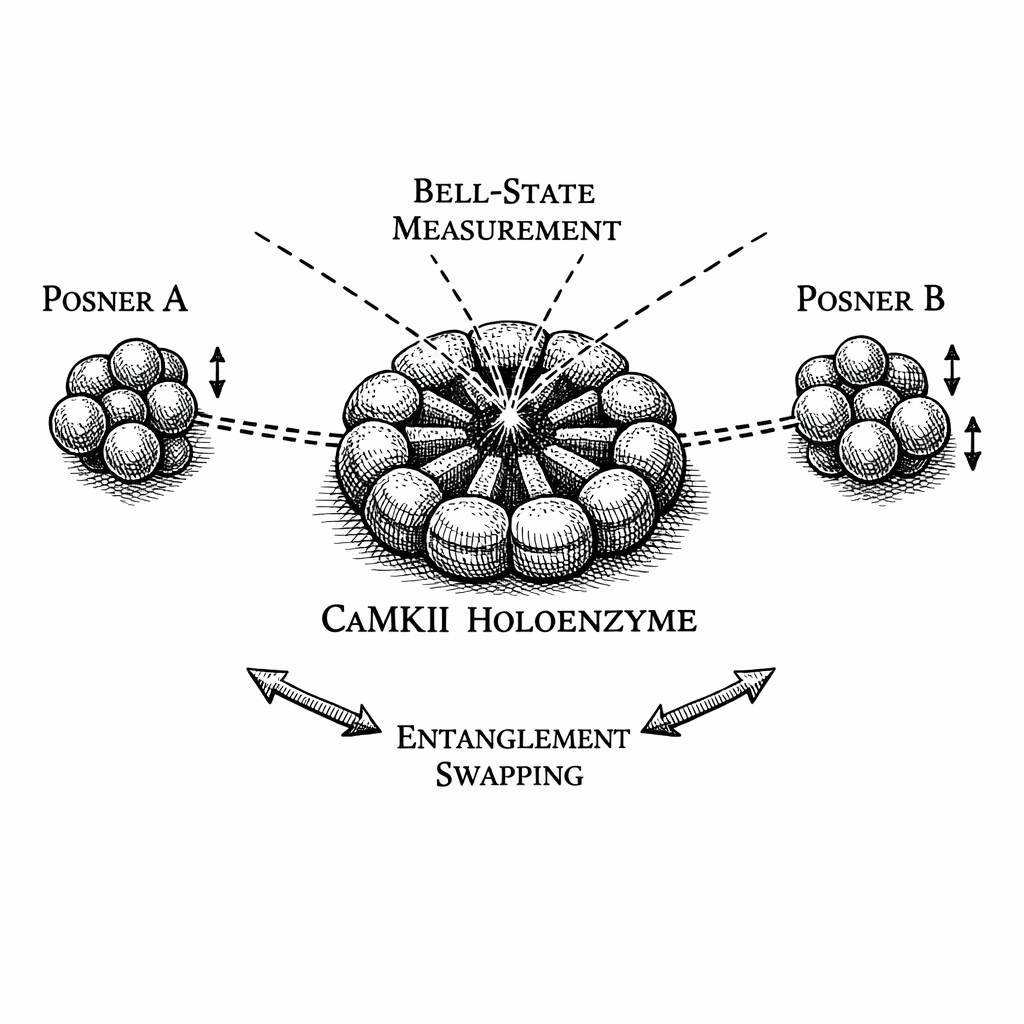
Now, suppose a researcher takes Particle B and Particle C, brings them together, and performs a joint quantum measurement known as a Bell-State Measurement.
The mathematics of quantum mechanics dictate a specific result. The moment B and C interact, their original entanglements to their partners are destroyed. Instantly and non-locally, Particle A and Particle D become quantum entangled with each other.
Particle A and Particle D have never physically touched. They have never shared a spatial location or interacted with the same environment. Yet, because their proxy partners interacted, A and D are now mathematically locked together, sharing a single wave function across macroscopic distances.
Entanglement can be passed down a chain like a virus.
Applying the mathematics of Entanglement Swapping to the biological architecture of the human brain:
Neuron 1 (in the visual cortex) generates an entangled pair of Posner molecules (A and B). Neuron 2 (in the auditory cortex) generates its own entangled pair of Posner molecules (C and D).
Neuron 1 fires an action potential, passing Posner B across a chain of synapses into a connected hub cell, Neuron 3 (perhaps located in the thalamus, a central relay station). Neuron 2 also fires, passing Posner C into that same hub cell.
Inside the dendritic spine of Neuron 3, Posner B and Posner C drift in the fluid. A random thermal collision in the cytoplasm would simply cause decoherence. To achieve Entanglement Swapping, quantum mechanics requires a structured operation: a Projective Bell-State Measurement.
The framework proposes it relies on the Post-Synaptic Density (PSD)---a lattice of scaffolding proteins located inside the receiving neuron. Specifically, the Posner molecules are drawn into the symmetric hexagonal rings of the CaMKII holoenzymes. This protein matrix acts as a biological Bell-State Analyzer.
When Posner B and Posner C are caught in adjacent, symmetrical binding pockets of the CaMKII lattice, the enzyme aligns them. It forces their electron clouds to overlap in a controlled geometry, allowing their magnetic fields to cross-correlate. The chemistry executes a joint quantum measurement on the two vaults without exposing them to the thermal bath.
The moment they interact, Entanglement Swapping is triggered.
Instantly and non-locally, Posner A (in the visual cortex) becomes quantum entangled with Posner D (in the auditory cortex).
The entanglement does not need to physically travel the distance. It cascades through the billions of synaptic connections of the brain---not at the speed of chemical diffusion, but instantaneously, riding the non-local geometry of the Einstein-Rosen wormholes in the Holographic Bulk detailed in Chapter 3.
As billions of neurons fire action potentials, they mix, swap, and distribute Posner molecules. The Entanglement Swapping protocol triggers a continuous chain reaction.
Within minutes of metabolic activity, the volume of the human brain---from the brainstem to the prefrontal cortex---is swept into a unified web of quantum entanglement.
14.6 Macroscopic Coherence: The Birth of the Antenna
Consider the architectural sequence biology has engineered.
We began in Chapter 11 with a single enzymatic cleavage of an ATP molecule, generating a pair of entangled phosphorus atoms.
By utilizing the calcium armor of the Posner molecules, the Trojan Horse mechanism of vesicular exocytosis, and the mathematics of Entanglement Swapping, biology scales that microscopic quantum state into a macroscopic reality.
The 86 billion isolated neurons are no longer classical islands. Beneath the electrical activity of the action potentials, the brain is united by a non-local web of entangled nuclear spins.
The brain has achieved Macroscopic Quantum Coherence.
The organ operates as a unified Decoherence-Free Subspace. The Posner molecules spread across the cortex are not individual quantum systems; they are a single quantum object. They share a brain-wide mathematical wave function ().
The framework provides a physical and mathematical resolution to the Binding Problem.
You do not experience the cardinal's color, its song, and its location as disjointed data streams because the neurons processing that data are quantum entangled. They are bound together into a single geometric state.
When the observer wave function () focuses its attention, generating a Fisher Information gradient in the Semantic Dimension, the thermodynamic force of that attention does not collapse a single atom.
Because the Posner network is entangled, the thermodynamic force of attention collapses the wave function of the brain simultaneously.
The macroscopic wave function collapses. The symmetry of the brain-wide Posner network is broken in a single, non-local instant. The visual, auditory, and cognitive realities snap into existence concurrently.
The Biological Antenna is now online. It is tuned to the 0.1 eV mass scale of the Semantic Dimension, secured within the Decoherence-Free Subspace.
The framework has modeled the receiver. We understand how the Semantic Dimension interacts with the signal. But we face a final mechanical gap in Dimensional Field Theory.
How does the collapse of a quantum state inside a calcium shell cause a human being to move their arm? How does the mind's thermodynamic push translate into a macroscopic electrical cascade? How does the observer pull the biological trigger?
To complete Part VI, we must map the chemical cascade that occurs the moment the wave function collapses. We must explore the mechanism of Downward Causation, and the Calcium Avalanche.
References - Chapter 14:
[1] Revonsuo, A. (1999). Binding and the phenomenal unity of consciousness. Consciousness and Cognition, 8(2), 173-185.
[2] Fisher, M. P. A. (2015). Quantum cognition: The possibility of processing with nuclear spins in the brain. Annals of Physics, 362, 593-602.
[3] Pan, J. W., Bouwmeester, D., Weinfurter, H., & Zeilinger, A. (1998). Experimental entanglement swapping: entangling photons that never interacted. Physical Review Letters, 80(18), 3891.
Discussion (0)
No comments yet. Start the conversation!