Part VI: The Biological Receiver
Chapter 15: The Calcium Avalanche (Downward Causation)
15.1 The Final Abyss: The Paradox of Downward Causation
In the philosophy of science and the standard model of modern physics, there is a concept known as Downward Causation. It is the proposition that a higher-level, macroscopic system (like a conscious mind, a social structure, or a global climate pattern) can reach "downward" and actively dictate the physical behavior of its fundamental, microscopic constituent parts (like individual cells or atoms).
For over a century, classical physics and biology have largely treated Downward Causation as a mathematical impossibility.
The classical worldview is reductionist and operates entirely via Upward Causation. It posits that the universe is built strictly from the bottom up. The fundamental properties of quarks determine the behavior of protons; the behavior of protons determines the chemistry of atoms; the interactions of atoms dictate the folding of proteins; the folding of proteins drives the firing of neurons; and the firing of neurons creates the emergent experience of a conscious mind.
In this paradigm, the mind is an "epiphenomenon"---the exhaust smoke rising from the mechanical engine of the brain. The smoke cannot reach down into the engine block and change the timing of the spark plugs. It possesses no causal power.
If Dimensional Field Theory (DFT) is to survive as a viable physical cosmology, it must address this reductionist model. We must demonstrate that the smoke turns the wheel.
We have spent three volumes building the architecture of the mind. We have established the Semantic Dimension. The framework proposes that the biological brain organically manufactures a macroscopic quantum network of entangled Phosphorus-31 nuclear spins, shielded inside millions of Posner molecules (). We have mathematically demonstrated that, within this framework, when the conscious observer () focuses its attention, it generates a thermodynamic Fisher Information gradient in the Holographic Bulk.
But a thermodynamic ripple in a higher dimension is functionally useless if it cannot move the heavy, wet machinery of the physical 3D Boundary.
We now face the final physical abyss.
When the thermodynamic force of human attention strikes the Decoherence-Free Subspace of the brain, it collapses the quantum wave function of the entangled nuclear spins. The microscopic, 0.1 eV ghost has touched the machine.
But how does the collapse of a tiny, invisible nuclear spin deep inside an atom force a macroscopic biological neuron to fire an electrical action potential?
An action potential is a large-scale classical event. It requires the synchronized movement of tens of thousands of heavy sodium and potassium ions across a lipid membrane, generating a voltage shift of roughly 100 millivolts. How does a mathematical anomaly translate into the high-voltage electrical storm required to move a human arm?
To a classical physicist, this is the equivalent of dropping a single grain of sand onto a continent and expecting it to trigger a magnitude 9.0 earthquake. The energy scales are mismatched by millions of orders of magnitude.
But this objection assumes that the biological brain is a passive, inert system. It is not.
The brain is a pressurized, non-equilibrium thermodynamic engine. It rests on a biological hair-trigger. To trigger an earthquake, you do not need to push the continental tectonic plates with your bare hands. You only need to apply a microscopic amount of friction to the exact fault line holding back the accumulated pressure of a billion tons of rock.
The biological fault line of the human brain is the Posner molecule. To understand how the ghost pulls the trigger, we must examine the structural tension holding this quantum vault together.
15.2 The Tensegrity of the Metastable Vault
To understand the mechanical bridge of the brain, we must look deeply into the physical chemistry of the biological quantum vault: the Posner molecule.
As we detailed in Chapter 11, the Posner molecule () is a highly symmetrical, roughly spherical cluster. Nine positively charged calcium ions () form an outer shell, encapsulating six negatively charged orthophosphate ions ().
In the language of thermodynamics, the Posner molecule is not a permanent, unbreakable rock. It is a metastable structure.
When bone mineralogists study calcium phosphate clusters, they typically observe them in the skeletal system. Here, these molecules rapidly aggregate, crystallize, and drop into their lowest energy state, forming a rigid mineral known as hydroxyapatite. Hydroxyapatite is thermodynamically stable. It is bone.
But when Posner molecules are formed in the fluid cytoplasm of a living neuron, the cell prevents them from crystallizing. The neuron surrounds them with specialized proteins and regulates the intracellular pH to keep the Posner molecules in a state known as Amorphous Calcium Phosphate (ACP).
A metastable amorphous structure is a physical system that is temporarily stable, but is hovering on the edge of a major energy transition. Imagine a heavy boulder resting in a shallow depression at the top of a steep mountain. As long as the environment is calm, the boulder will sit there indefinitely. It appears stable. But it only takes a microscopic push---the weight of a single bird landing on it, or a slight vibration in the rock---to roll it out of the depression. Once it breaches that lip, gravity takes over. The boulder accelerates down the mountain, releasing a large amount of stored kinetic energy.
The Posner molecule is the biochemical equivalent of that boulder.
It relies on a delicate biological "tensegrity" (tensional integrity). The nine calcium ions (totaling a +18 charge) and the six phosphate ions (totaling a charge) are locked together by sensitive electrostatic bonds. The molecule maintains its spherical geometry only because the repulsive forces between the highly negative phosphates in the core are counterbalanced by the attractive, constricting forces of the positive calcium shell.
The molecule is a pressurized container. It holds pent-up electromagnetic potential, balanced on a razor's edge.
It maintains its structural integrity against the thermal bombardment of the 300-Kelvin cytoplasm through its spherical symmetry. As long as the geometric symmetry is preserved, the forces cancel out, and the vault survives.
It is a structure waiting for a microscopic push.
15.3 The Collapse and the Hyperfine Fracture
Let us slow time down to the scale of femtoseconds ( seconds), and observe the exact moment of Downward Causation.
Across the volume of the human cortex, millions of Posner molecules float in the intracellular fluid and synaptic clefts. Because of the mathematics of Entanglement Swapping (detailed in Chapter 14), these molecules share a single, macroscopic quantum wave function. The Phosphorus-31 nuclei inside them are locked in a balanced Singlet State (Total Spin = 0). They exist in a quantum superposition. Their magnetic fields are blurred into a symmetrical haze.
The conscious observer makes a choice.
In the Semantic Dimension, the observer wave function () narrows its focus. The internal informational entropy of the mind drops. A Fisher Information gradient forms in the Holographic Bulk.
The universe, naturally opposing this thermodynamic gradient, seeks to balance the ledger. Entropic Gravity transmits this tension across the dimensional boundary.
The thermodynamic force strikes the Decoherence-Free Subspace of the brain. Because the Posner network is isolated from classical noise and tuned to the 0.1 eV mass scale of the Semantic Field, the force encounters no classical resistance.
The thermodynamic tension shatters the superposition. The macroscopic wave function of the brain collapses.
Inside the Posner molecules, the unresolved Singlet State of the phosphorus nuclei is broken. The wave function collapses into a definite physical state. The nuclear spins snap into alignment (e.g., strictly Spin-Up or Spin-Down). The quantum ghost has become a physical reality.
But what happens to an atom when its nucleus snaps from a spherical superposition into a definite, directional state?
In basic chemistry, the nucleus of an atom is frequently treated as a static weight, while chemical bonding is handled exclusively by the orbiting electron cloud. But in quantum chemistry, the nucleus and the electrons engage in a constant magnetic dialogue. This bridge is known as the Hyperfine Interaction.
The Hyperfine Interaction is the magnetic coupling between the magnetic dipole moment of the spinning nucleus and the magnetic field generated by the surrounding electrons.
When the nuclear spin of the Phosphorus-31 atom collapses into a definite alignment, a highly localized magnetic field flares into existence at the center of the atom.
This magnetic shift propagates outward through the Hyperfine Interaction, pulling on the surrounding electron cloud of the phosphate ion.
The electrons act as the glue holding the Posner molecule together. When the electron cloud of the central phosphorus atoms shifts, it is microscopically warped. The electron orbitals are pulled out of spherical alignment.
The spherical symmetry of the quantum vault is broken.
(Note: The Hyperfine Interaction is well-documented in biology. It is the quantum mechanism that allows migratory birds, like the European Robin, to navigate. The bird's retina contains a protein called cryptochrome, which forms entangled radical pairs. The Earth's weak magnetic field alters the nuclear spins of these atoms. Through the Hyperfine Interaction, this alters the electron chemistry, allowing the bird to "see" the magnetic field of the Earth. Evolution is already familiar with spin chemistry.)
In the metastable architecture of the Posner molecule, this electron warp is the equivalent of the bird landing on the boulder. It is the exact "activation energy" required to destabilize the structure.
A geometric fault line propagates through the electrostatic bonds. The thermodynamic ripple from the Semantic Dimension provides the infinitesimal push required to breach the lip of the mountain.
15.4 Maxwell's Demon and the Crystalline Precipice
At this juncture, a classical thermodynamicist might raise a significant objection based on the Boltzmann constant (). In the 300-Kelvin environment of the human brain, the ambient thermal noise---the baseline kinetic energy of molecules vibrating due to heat---is roughly electron volts (eV).
If the thermodynamic force of the conscious mind is mediated by a coupling constant of , how can a force that infinitesimally weak overcome an activation barrier against the eV thermal background of biology? Furthermore, if the non-physical mind does inject enough raw kinetic energy to break a molecular bond, hasn't Dimensional Field Theory just violated the First Law of Thermodynamics (Conservation of Energy), committing the same error that constrained Rene Descartes four centuries ago?
To address this, we must look to non-equilibrium statistical mechanics, and revisit a paradox from 1867: Maxwell's Demon.
First, the biological brain is not a passive system waiting to be pushed. It is a non-equilibrium engine. The brain continuously burns metabolic energy (via the hydrolysis of billions of ATP molecules) to push the Posner molecule network to the edge of a phase transition.
In statistical mechanics, this state is known as Self-Organized Criticality (SOC). When a physical system is held at a critical phase transition---like water suspended just before freezing into ice, or a pile of sand just before an avalanche---it develops what physicists call infinite susceptibility. The correlation length of its internal fluctuations diverges. In this primed state, the system does not require a high-energy impact to collapse. The macroscopic kinetic energy required to shatter the vault was already paid for by the biology using ATP. The biological spring is coiled; the energy barrier effectively drops to zero.
But who pulls the trigger? And how is it pulled without expending physical energy?
In 1867, James Clerk Maxwell imagined a microscopic demon sitting at a door between two chambers of gas. By observing the speed of the gas molecules and opening or closing the door at the right time, the demon could sort the hot molecules from the cold ones, decreasing entropy and doing physical work without expending kinetic energy to push the molecules. For decades, this threatened to break the Second Law of Thermodynamics.
In 1929, the physicist Leo Szilard solved the paradox. He proved mathematically that the demon does pay a thermodynamic toll, but it pays it not with kinetic energy, but with Information. The act of measuring the particles, storing that information, and then erasing the demon's memory generates the entropy required to balance the cosmic ledger. Szilard proved that Information and Energy are interchangeable (). A "Szilard Engine" mathematically converts information into physical work.
In Dimensional Field Theory, the conscious observer () operating in the Semantic Bulk operates as the physical realization of Maxwell's Demon.
When the mind focuses its attention, it does not inject physical kinetic energy into the brain. It injects Information (negentropy). Because the wave function of the Posner network exists in a state of quantum probability, the observer's attention executes a unitary phase shift. It performs a topological rotation of the probability amplitude. This geometric rotation requires exactly zero kinetic energy, preserving the First Law of Thermodynamics.
The ripple provides the informational key that unlocks the infinite susceptibility of the critical state. The electrostatic bonds fail, the tensegrity breaks, and the pre-existing biological energy of the vault unwinds. Mind moves matter by balancing the thermodynamic ledger of the Szilard Engine.
15.5 The Calcium Avalanche
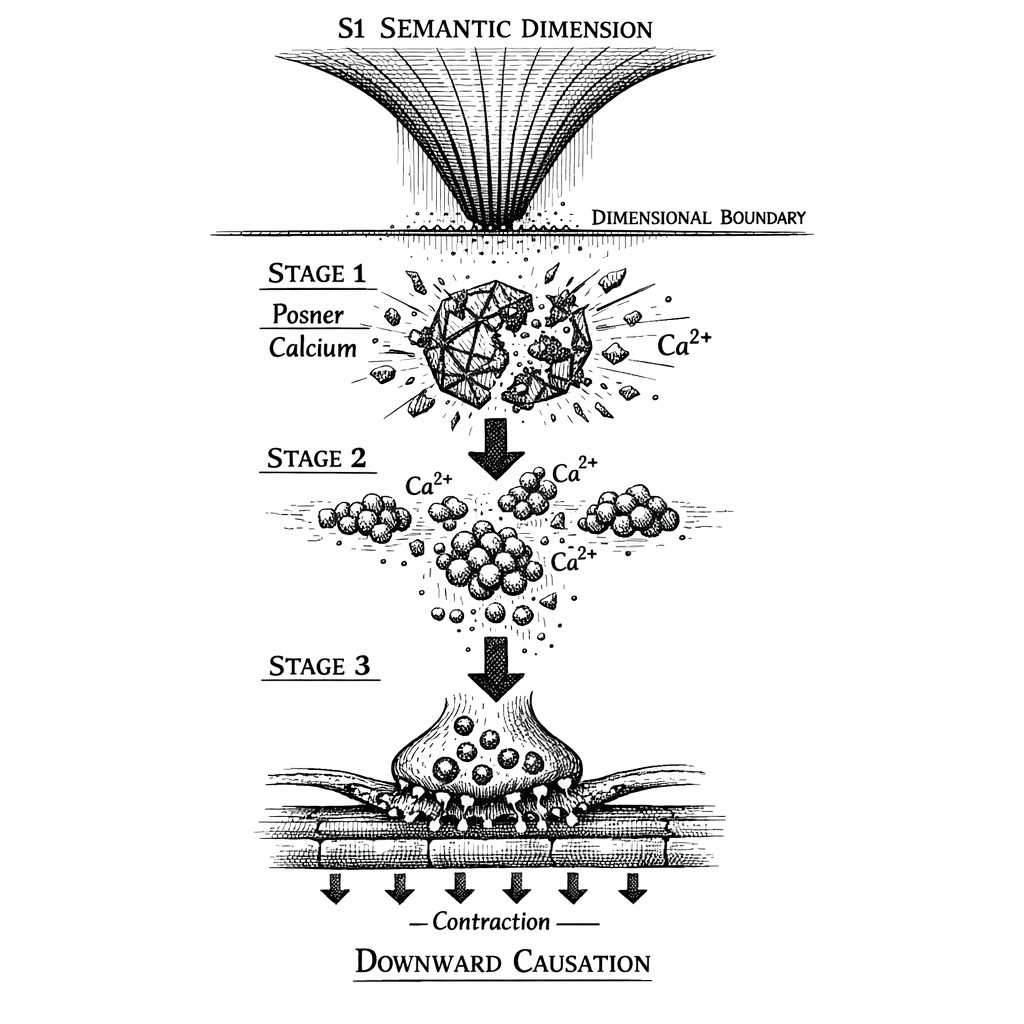
The moment the geometric symmetry is broken, the Posner molecule can no longer hold itself together against the thermal bombardment of the cytoplasm.
The electrostatic bonds shatter. The repulsive forces of the negatively charged phosphates inside the core overpower the weakened calcium shell. The structural integrity of the biological quantum vault fails.
The Posner molecule breaks apart, releasing its nine bound calcium ions () into the intracellular fluid.
To a classical biologist, nine calcium ions are insignificant. They are insufficient to bind to enough Synaptotagmin proteins to trigger vesicular release. But biology relies on cascade amplification. The Posner molecule does not act as the flood; it acts as the detonator.
Posner molecules do not float blindly in the void; they are electrostatically docked to the allosteric binding sites of classical protein floodgates known as Ryanodine Receptors (RyRs). These receptors sit on the surface of the Endoplasmic Reticulum (ER)---the neuron's internal storage organelle, holding millions of calcium ions.
When the quantum vault shatters, the localized spike of nine calcium ions occurs at point-blank range to the Ryanodine Receptor. This triggers a classical biological mechanism known as Calcium-Induced Calcium Release (CICR).
The Ryanodine Receptor detects the micro-burst, its floodgates open, and the ER releases its payload. Millions of stored calcium ions pour out of the Endoplasmic Reticulum and flood the synapse. The microscopic quantum spark has ignited the classical powder keg.
Because the Posner network was quantum entangled, this shattering does not happen in a single, isolated molecule. When the macroscopic wave function collapsed, it collapsed the entire network. Across the volume of the targeted neural pathways---in the prefrontal cortex, the supplementary motor area, and the visual cortex---millions of Posner molecules shatter in the same microsecond, each detonating its own local Ryanodine cascade.
We have achieved the true Calcium Avalanche.
To a quantum physicist, the collapse of the wave function is complete. The Semantic Dimension has finished its work. But to a neurobiologist, the cascade has just begun.
15.6 The Classical Ignition (Synaptotagmin and Calmodulin)
In classical neuroscience, one principle is broadly accepted: Intracellular Calcium is the master ignition switch of the brain.
Under resting conditions, a neuron expends ATP energy to pump free calcium out of its cytoplasm. It maintains an internal calcium concentration roughly 10,000 times lower than the fluid outside the cell. The neuron keeps its interior starved of calcium so that when a calcium flood does occur, the cell reacts with high-priority speed.
When the millions of Posner molecules shatter, they bypass the neuron's guarded external calcium channels. The resulting Calcium Avalanche creates localized spikes of calcium concentration---known in neuroscience as Calcium Micro-domains---deep inside the neurons and directly at the axon terminals.
These free calcium ions are instantly snatched up by highly sensitive calcium-binding proteins.
1. The Synaptic Trigger (Synaptotagmin):
At the axon terminal, the sudden flood of calcium binds to a specialized sensor protein called Synaptotagmin-1 [1]. As explored in Chapter 14, Synaptotagmin is the biological detonator for neurotransmitter release. When the calcium binds to its C2A and C2B domains, the protein changes its physical shape. It plunges into the lipid membrane, interacting with the SNARE protein complex and forcing it to rapidly zipper together.
This mechanical zippering forces the synaptic vesicles to fuse with the cell membrane. The vesicles burst open, releasing classical neurotransmitters (like glutamate or dopamine) into the synaptic cleft. The neurotransmitters bind to the neighboring neuron, sodium channels snap open, and a 100-millivolt electrical action potential begins. The neuron has successfully fired a macroscopic signal.
2. The Memory Forge (Calmodulin):
Simultaneously, deeper inside the body of the cell (the soma) and the dendritic spines, the calcium binds to a messenger protein called Calmodulin [2].
When activated by the calcium avalanche, Calmodulin acts as a biological master key. It unlocks cascades of kinase enzymes (specifically CaMKII). These enzymes travel to the surface of the neuron and alter the structure of the AMPA receptors, keeping them open to allow more electrical charge to flow in the future. Furthermore, they travel into the nucleus of the cell, altering the transcription of DNA to build new synaptic connections.
This is the biochemical mechanism of Long-Term Potentiation (LTP) [3].
The mind has not just fired the neuron; by triggering the calcium-calmodulin cascade, the mind has physically rewired the hardware of the brain. The non-physical thought has forged a permanent physical memory of the conscious experience.
15.7 The Unbroken Chain (Closing the Cartesian Divide)
Consider the unbroken chain of physical causality we have mapped across three volumes of physics and biology.
We have constructed a mathematically explicit and biochemically grounded bridge across the Cartesian Divide. The framework maps the sequence of events from philosophical intention to the electrical reality of moving muscle.
The Semantic Choice: The human observer, existing as a geometric information structure in the Holographic Boundary, chooses to focus their attention.
The Thermodynamic Gravity: This psychological act of focus decreases the observer's informational entropy, generating a Fisher Information gradient in the Semantic Bulk.
The Entropic Force: To neutralize this thermodynamic gradient and balance the cosmic ledger, the universe applies a entropic force to the physical wave function on the Boundary.
The Quantum Collapse: This semantic force acts non-locally on the macroscopic, entangled Decoherence-Free Subspace of the brain's Posner molecule network, forcing the entangled Phosphorus-31 nuclear spins to collapse out of the Singlet State.
The Hyperfine Fracture: The nuclei snap into a definite spin state. This alters their magnetic fields, which warps the surrounding electron clouds via the Hyperfine Interaction, breaking the spherical symmetry of their molecular cages.
The Calcium Avalanche: The loss of symmetry acts as the activation energy that destabilizes the metastable electrostatic bonds of the Posner molecules. Millions of Posner vaults shatter simultaneously across the targeted brain regions, releasing a coordinated flood of intracellular calcium.
The Classical Ignition: The calcium micro-domains bind to Synaptotagmin and Calmodulin, triggering vesicular exocytosis and neuroplastic rewiring.
The Macroscopic Reality: The neurotransmitters bridge the synaptic cleft, opening sodium channels, and firing the electrical action potential that travels down the spinal cord. Muscle fibers contract. A human arm moves.
The ghost has moved the machine.
It did not do so by violating the First Law of Thermodynamics. It did not magically inject new kinetic energy into a closed universe. It utilized the thermodynamic laws of information geometry to collapse a primed, metastable quantum fault line, releasing a reserve of pre-existing biological energy.
The architecture of the biological antenna is complete. Part VI is resolved. We have mapped the physics, and we have mapped the flesh. Dimensional Field Theory offers a biologically viable cosmology.
But a theoretical framework cannot simply end with a biological schematic. A theory that models the fundamental architecture of the universe must also explain the human experience of that universe.
If this theory holds, it shifts our understanding of psychology, mental illness, the flow of time, and the concept of free will. We must take this mathematical and biological framework and translate it back into the reality of human existence.
What does this proposed cosmology mean for you, the conscious observer? How does the Fisher Information gradient map to the psychology of trauma, the focus of an athletic "Flow State," or the neurological chaos of Schizophrenia? What does it mean for the illusion of linear time, human regret, and the ultimate responsibility of true free will?
We must take the machinery of Parts V and VI, and build the Rosetta Stone. We must deconstruct the equations for humanity.
We open Part VII.
References - Chapter 15:
[1] Sudhof, T. C. (2012). The presynaptic active zone. Neuron, 75(1), 11-25.
[2] Clapham, D. E. (2007). Calcium signaling. Cell, 131(6), 1047-1058.
[3] Kauer, J. A., & Malenka, R. C. (2007). Synaptic plasticity and addiction. Nature Reviews Neuroscience, 8(11), 844-858.
Discussion (0)
No comments yet. Start the conversation!