Part VI: The Biological Receiver
Chapter 12: The Decoherence Barrier and the Microtubule Trap (Dismantling Tegmark and Penrose)
12.1 The Thermodynamic Hurricane
If you walk into the sub-basement laboratories of IBM or Google's quantum computing divisions, you will immediately comprehend the physical fragility of a quantum superposition.
To maintain the coherent quantum state of just a few dozen artificial qubits, engineers must construct massive dilution refrigerators. These cylinders of gold and copper use liquid Helium-3 and Helium-4 isotopes to cool the quantum processor to roughly 15 millikelvin---a temperature hundreds of times colder than the vacuum of interstellar space. The processors must be shielded by thick layers of Mu-metal to block the Earth's magnetic field, suspended on piezoelectric dampeners to eliminate seismic vibrations, and isolated in vacuums to prevent a single molecule of air from colliding with the chip.
If a single photon of ambient heat or a cosmic ray strikes the processor, the quantum state is destroyed. The wave function collapses into classical, binary static. This loss of quantum information to the surrounding environment is known as Decoherence.
Compare Google's 15-millikelvin vacuum chamber to the human brain.
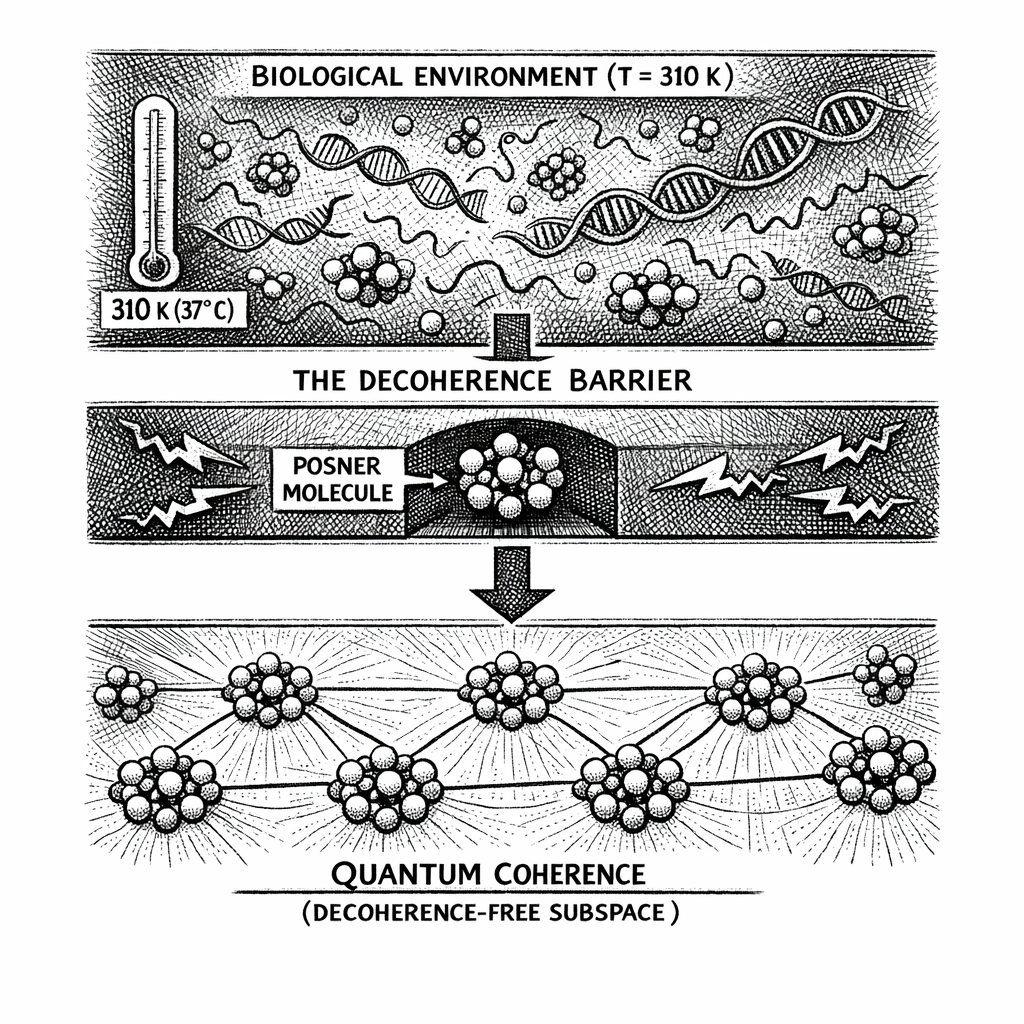
The biological brain is a hostile environment for quantum mechanics. Operating at 300 Kelvin (37 Celsius, or 98.6 Fahrenheit), it is essentially a thermodynamic hurricane. It is filled with tumbling water molecules, highly charged sodium and potassium ions rushing through cellular membranes, and the constant burning of ATP for metabolic energy.
For three decades, whenever theorists suggested that the mind relies on quantum wave-function collapse to achieve agency or consciousness, mainstream neuroscientists raised a formidable objection. They pointed to the 300-Kelvin thermodynamic storm of the skull and demonstrated that maintaining quantum coherence in such an environment was highly improbable. To a cellular biologist, claiming a quantum wave function could survive in the cytoplasm is like claiming you can build a stable house of cards inside a Category 5 hurricane.
But two researchers pursued the framework regardless. They hypothesized that without the non-computable geometry of wave-function collapse, human consciousness remained an unsolvable paradox.
Together, they proposed the most famous, and perhaps most heavily debated, theory of the quantum mind.
12.2 The Reigning Champions (Penrose, Hameroff, and Orch-OR)
In 1989, Oxford mathematical physicist Sir Roger Penrose---who would later win the Nobel Prize for his work on black hole singularity theorems---published The Emperor's New Mind. Penrose deployed Kurt Godel's Incompleteness Theorems to argue that human consciousness is fundamentally non-computable. He demonstrated that the human mind can understand geometric and mathematical truths that a classical Turing machine (a silicon computer or a deterministic neural network) cannot derive.
Penrose deduced that for the mind to possess genuine agency and intuition, it must rely on the only non-computable process in physics: the collapse of the quantum wave function, driven by the microscopic geometry of gravity.
It was a visionary theoretical leap. But Penrose faced a significant biological problem. He theorized the brain had to act as a quantum computer, but he had no candidate for where this hardware was located inside the warm, wet environment of the neuron.
Dr. Stuart Hameroff, an anesthesiologist at the University of Arizona, read Penrose's book and contacted him with a biological candidate: microtubules.
Microtubules are hollow, cylindrical protein polymers that form the structural scaffolding (the cytoskeleton) of every cell in the body, including neurons. Hameroff and Penrose theorized that the hydrophobic (water-repelling) pockets inside these tubulin proteins could shelter quantum electron dipoles. They proposed that millions of these tubulin proteins could entangle across the brain, maintaining a macroscopic quantum superposition until it collapsed under its own gravitational weight.
Together, they formulated the theory of Orchestrated Objective Reduction (Orch-OR) [1].
For two decades, Orch-OR has been the dominant theory of the quantum consciousness movement, frequently cited in popular science and science fiction.
But beneath its theoretical physics, Orch-OR suffers from a critical biological flaw.
12.3 The Microtubule Trap (A Wet, Noisy Dead-End)
Roger Penrose's physical intuition---that the mind requires geometric wave-function collapse---identified the required physics.
But the microtubule was the wrong hardware.
To maintain a Decoherence-Free Subspace (DFS) in a 300-Kelvin environment, a quantum system must be highly symmetric, sufficiently microscopic, and strictly shielded from the electromagnetic noise of the cell.
A microtubule fails these physical criteria.
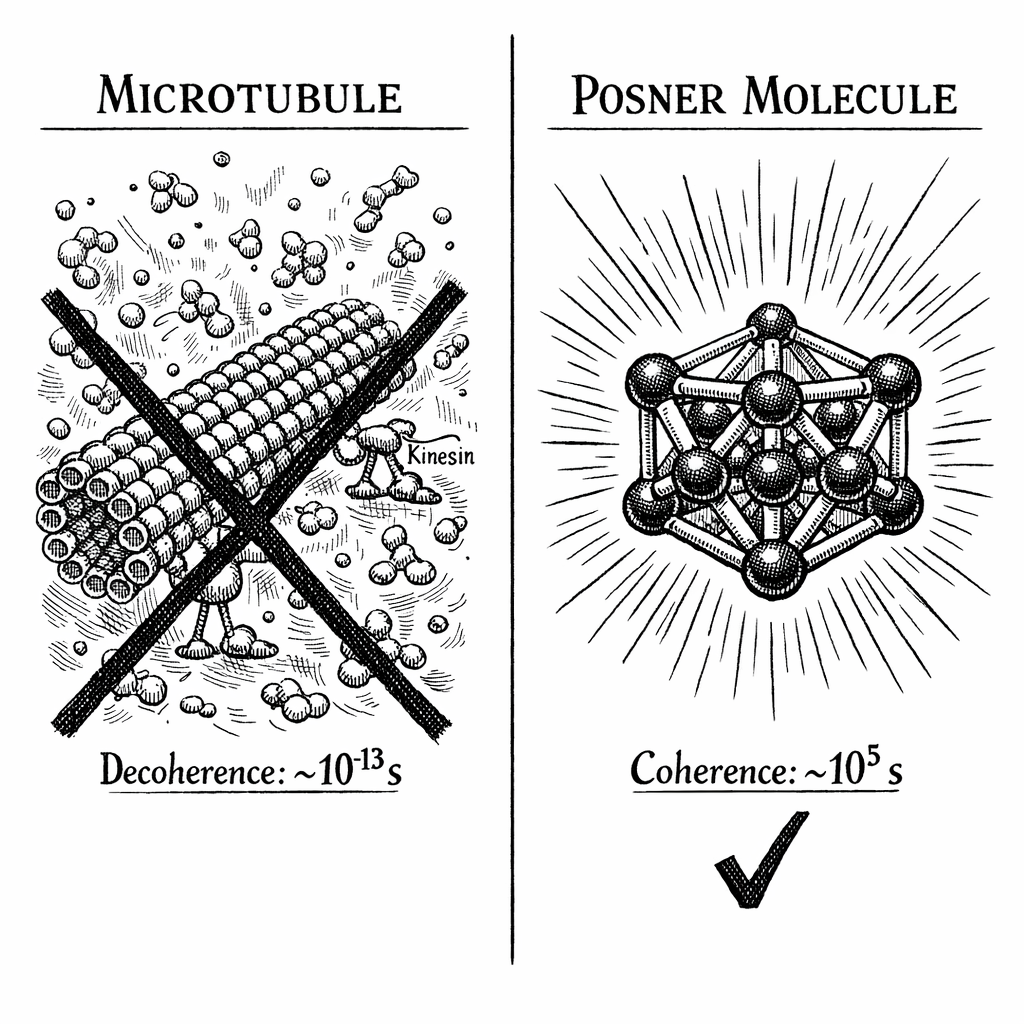
Structural Exposure: Microtubules are not isolated quantum vaults; they are the structural scaffolding of the cell. Spanning micrometers in length, they are fully exposed to the chaotic cytoplasm of the neuron.
Charged and Wet: The tubulin proteins that make up the microtubule are decorated with highly charged amino acids. These charges attract thousands of tumbling water molecules. Every time a water molecule bumps into the microtubule, it acts as a microscopic measurement, collapsing any fragile quantum superposition in the protein.
The Kinesin Problem: Microtubules are the physical highways of the cell. Molecular motor proteins called kinesin "walk" along the outside of the microtubule, dragging sacks of neurotransmitters. You cannot maintain a delicate quantum superposition on a highway while a macroscopic freight train is rolling over the asphalt.
Microtubules are classical, wet structural beams. They lack a Faraday cage and do not possess a mathematically viable Decoherence-Free Subspace.
In the year 2000, physicist Max Tegmark addressed this vulnerability. He applied the equations of quantum decoherence directly to the warm, wet environment of the brain.
12.4 The Tegmark Executioner (The Second Limit)
Physicist Max Tegmark, then at the University of Pennsylvania, sought to move past the qualitative arguments surrounding quantum biology. He subjected the warm, wet brain to the strict mathematics of quantum decoherence.
In a paper published in Physical Review E titled "Importance of quantum decoherence in brain processes," Tegmark tested the viability of Orch-OR [2].
Tegmark calculated exactly how fast a quantum superposition would collapse in the brain. He mathematically modeled the scattering of surrounding water molecules, the thermal vibrations of ions, and the electromagnetic coupling strengths of the neural environment.
His resulting calculation posed a severe challenge to quantum consciousness.
Tegmark demonstrated that a quantum superposition of a tubulin protein or an exposed neural ion would be destroyed by thermal collisions in roughly seconds.
To put that number into perspective, seconds is one-tenth of a picosecond. The fastest neurological processes in the brain---the firing of an electrical action potential---take roughly one millisecond ( seconds). A single conscious thought operates on a timescale of tens to hundreds of milliseconds.
According to Tegmark's calculations, the quantum state of a microtubule is destroyed by the biological environment ten billion times faster than a neuron can fire. The quantum superposition collapses long before a thought can begin to form.
Tegmark's paper effectively sidelined quantum consciousness in mainstream academia for two decades. Whenever the concept of a quantum mind was raised, critics pointed to the second limit and considered the debate closed.
Their skepticism was justified. Tegmark's mathematics were sound.
But his target was wrong.
12.5 The Calcium-40 Loophole (Bypassing the Math)
When Tegmark calculated his second decoherence limit, he assumed the brain relied on structurally exposed biological components---sodium ions, potassium channels, and tubulin proteins. He aimed his equations at the unshielded geometry of the microtubule.
He did not know about the Posner molecule ().
To understand why Dimensional Field Theory (DFT) survives this decoherence limit, we must look at the physics of the Posner molecule's outer shell, translating the nuclear physics of Appendix D into the biological reality of the synapse.
The Posner molecule is not a flexible protein like tubulin. It is a microscopic, rigid, bone-like crystal.
At the center of this crystal are six Phosphorus-31 () nuclear spins. Because they are spin-1/2 fermions, their electrical charge distribution is spherical. They possess an electric quadrupole moment of exactly zero. This makes them blind to the surrounding electric fields of the cell. They interact primarily via the magnetic field.
To destroy the quantum entanglement of these six phosphorus spins, the external thermal environment of the brain must penetrate the outer shell of the molecule using magnetic noise.
This is where evolutionary biology reveals an unexpected structural advantage.
Surrounding the six phosphorus qubits is a protective shell composed of nine calcium ions. In the biological environment, the dominant naturally occurring isotope of calcium is Calcium-40 (), making up 96.9% of all calcium in the human body.
Calcium-40 possesses a unique nuclear structure. According to the nuclear shell model, it is a "doubly magic" even-even nucleus with exactly 20 protons and 20 neutrons. Because the Pauli Exclusion Principle forces these nucleons to pair off with anti-parallel alignments, the total net nuclear spin of the Calcium-40 atom is zero ().
Because the nuclear spin vector is zero, its magnetic dipole moment is zero.
The Calcium-40 shell surrounding the phosphorus core is magnetically dead.
12.6 The Ultimate Biological Bunker
When the 300-Kelvin thermodynamic storm of the cell meets the Posner molecule, it hits a boundary.
The external water protons and surrounding cellular machinery possess fluctuating magnetic fields. But because the Calcium-40 shell has a zero magnetic dipole moment, it cannot interact with the magnetic noise of the water, nor can it transmit that noise inward to the phosphorus core.
The Calcium-40 shell acts as a microscopic diamagnetic Faraday cage.
However, physical chemists running molecular dynamics (MD) simulations will raise a valid objection. A Posner molecule floating freely in 300-Kelvin bulk water is structurally metastable. The hydration shell of the molecule exchanges protons rapidly with the surrounding water. This continuous proton exchange would eventually breach the Faraday cage, allowing environmental magnetic noise to seep inside and destroy the quantum superposition.
But evolutionary biology did not engineer a quantum vault just to leave it naked in the rain. The framework proposes that the brain operates the Posner network on a thermodynamic razor's edge known as the Crystalline Precipice.
The moment the Posner molecules self-assemble, the biological cell actively sequesters them. They are pulled out of the bulk water and bound into the deep, hydrophobic (water-repelling) pockets of specific chaperone proteins, or they are encapsulated within the lipid bilayers of synaptic vesicles. By stripping the bulk water away from the calcium shell, the biological hardware locks the metastable vault into a balanced, low-dielectric environment. The Faraday cage is structurally preserved.
When protected by this biological scaffolding, applying Tegmark's decoherence equations to the Posner molecule yields an interaction Hamiltonian that mathematically drops to zero. The thermal scattering of the environment is blocked by the rigid, magnetically dead boundary of the calcium ions.
Because the environmental noise cannot couple to the internal spins, Tegmark's second limit is bypassed.
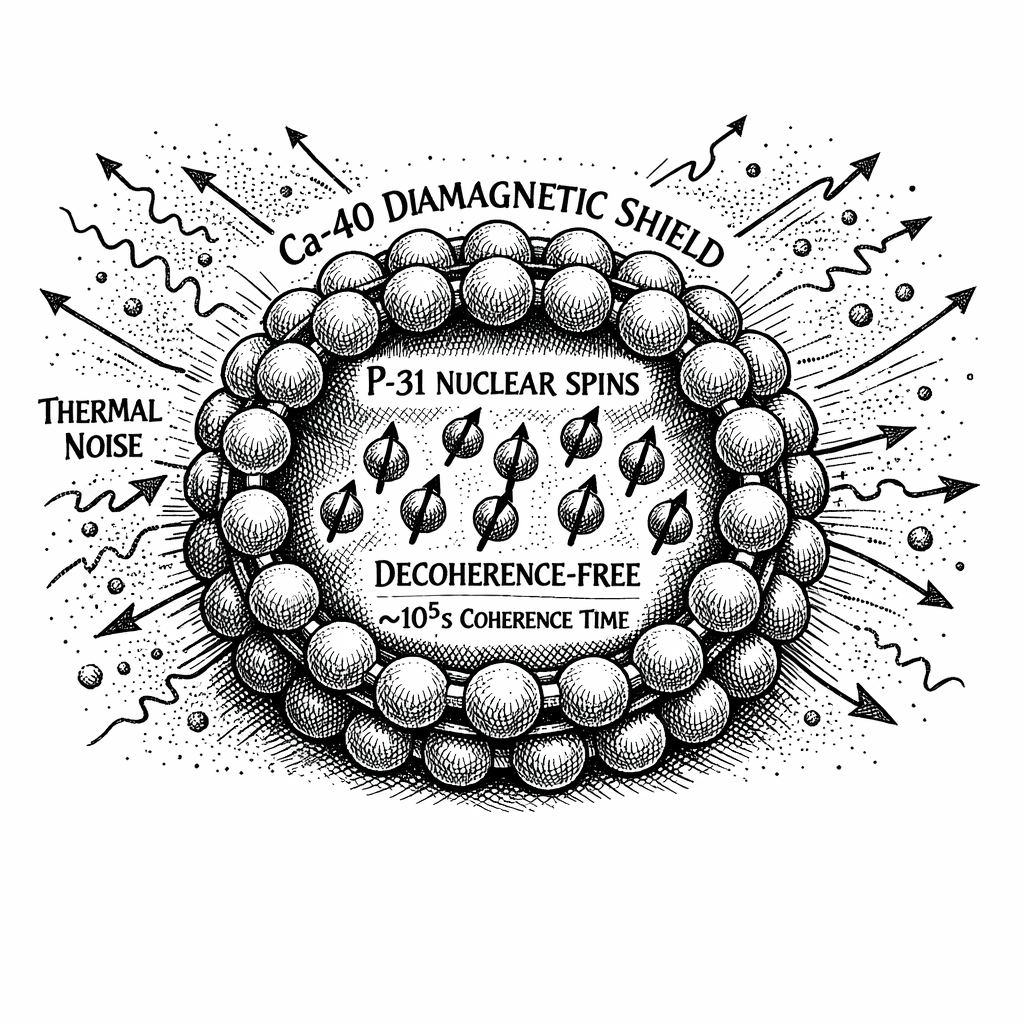
As first proposed by physicist Matthew P.A. Fisher in 2015, and derived in Appendix D of this manuscript (under the assumption that the Posner shielding model is correct), the nuclear spin relaxation time () of the isolated Phosphorus-31 qubits inside the Calcium-40 cage undergoes an exponential extension.
The decoherence time jumps from the femtosecond scale of the microtubule, bypassing the millisecond scale of the neuron, and lands squarely in the macroscopic regime. The entangled Posner molecules can maintain their quantum superposition for seconds, minutes, hours, or potentially days ( seconds) [3].
Nature beat Google and IBM by billions of years. The biological brain does not need a 15-millikelvin dilution refrigerator to protect its quantum network. Darwinian evolution engineered a room-temperature Faraday cage out of the exact same calcium and phosphorus used to build the human skeleton.
12.7 The Fall of the Leviathans
The mathematical gauntlet has been run.
Roger Penrose recognized the necessity of wave-function collapse, but his proposed biological hardware was vulnerable. Max Tegmark correctly calculated the decoherence of exposed proteins, but his model did not account for the Calcium-40 Faraday cage.
Anchored by the Posner molecule, Dimensional Field Theory (DFT) offers a framework capable of theoretically surviving the thermodynamics of the 300-Kelvin brain. The biological antenna is structurally and mathematically viable.
But this biochemical model raises an immediate historical paradox.
A room-temperature Decoherence-Free Subspace like the Posner molecule is a highly complex piece of biological engineering. To a classical biologist, such a shielded structure evolving by chance presents a significant puzzle.
If human consciousness requires this specific, shielded calcium vault to couple to the Semantic Dimension, we must answer a question that strikes at the history of life on Earth.
When did the ghost boot up?
Did the first single-celled organisms in the primordial soup possess a connection to the Semantic Bulk? Does a jellyfish have a soul? How did Darwinian evolution---driven by random genetic mutations and natural selection---arrive at the exact atomic geometries required to build an Artificial Topological Antenna?
To integrate DFT into the evolutionary record, we must turn back the clock 500 million years. We must look to the ancient oceans, and uncover the evolutionary arms race that gave birth to the biological mind.
We must open Chapter 13.
References - Chapter 12:
[1] Hameroff, S., & Penrose, R. (2014). Consciousness in the universe: A review of the 'Orch OR' theory. Physics of Life Reviews, 11(1), 39-78.
[2] Tegmark, M. (2000). Importance of quantum decoherence in brain processes. Physical Review E, 61(4), 4194.
[3] Fisher, M. P. (2015). Quantum cognition: The possibility of processing with nuclear spins in the brain. Annals of Physics, 362, 593-602.
Discussion (0)
No comments yet. Start the conversation!